Golgi apparatus is ubiquitous in cells. They are found in living cells of both animals and plants and are a dynamic structure. In animals, Golgi bodies usually appear morphologically as a cluster in one part of the cell and are interconnected with one another through tubules like a fibrous network. But, in plants, a large number of Golgi bodies are dispersed throughout the cytoplasm as separate elements. Golgi bodies are observed abundance in most secretory cells. However, the Golgi complex is not found in bacteria and blue-green algae and may be absent in dry seeds.
Discovery

Camillo Golgi, an Italian neurologist, first discovered these organelles with a light microscope in the Purkinje cells of the owl, Strix flammea. His staining method for light microscopy specimens pioneered the study of nerve cells to be crucial for the invention of the “internal reticular apparatus” in 1898. The Golgi complex, so named after its discoverer is a featured system of cytoplasmic membranes.
The membranes of the Golgi are discontinuous, smaller, and more compact in nature. It is still controversial that there is a direct morphological connection between the endoplasmic reticulum and the Golgi complex. However, it was early identified by its affinity for osmium of silver-containing stains. In modern days its existence has been explicitly visualized with the electron microscope.
Various workers have given different kinds of names to this complex, such as dictyosome, idiosome, lipochondria, Golgi body, Golgi substance, Golgi apparatus, and Golgi complex. Generally, the name Golgi complex deals with the material in vertebrates and dictyosome in invertebrates and plants.
Structure
There is variable size of Golgi complex. It is small in muscle cells but quite big in the nerve and gland cells. Generally, the complex is well developed in an active cell on the other hand in old cells the complex diminishes in size and ultimately disappears. The electron-microscopic examination of thin sections of cells exhibits the Golgi apparatus as a characteristically structured system of smooth double membranes (sacculi), which are either settled in parallel stacks or are layered around vacuoles.
The Golgi complex comprises of one flattened sacs or cisternae, large clear vacuoles, and clusters of dense vesicles.
1. Flattened sacs or Cisternae
These are known as cisternae. The basic structural unit of the Golgi apparatus, however, is the Golgi cisterna.
- Those are slightly curved and are dilated (swollen) at the margins like flasks.
- Each Golgi stack is a flattened disc about 1 µm in diameter and 0.25 µm long.
- A Golgi sack typically consists of a stack of 4-7 flattened membranes or cisternae, although more than 20 cisternae may be present.
- In many algal cells, Golgi bodies are known to consist of up to 20 – 30 cisternae.
- Not all Golgi apparatus have their cisternae in stacks. The Golgi apparatus in the red alga, Cyanidioschyzon merolae, composes of only one or two cisternae. Likewise, in the cells of many filamentous fungi, the Golgi apparatus demonstrates as a single tubule known as a Golgi equivalent. The Golgi apparatus of the budding yeast Pichia comprises four cisternae.
- These structures are similar to the smooth surface endoplasmic reticulum and seem in sectional view as dense parallel membranes. The packed cisternae are often arrayed concentrically, enclosing regions of cytoplasm filled with numerous large vesicles.
- The membranes do not communicate with each
- Each cisterna is separated from the others in the stack by a minimal space of 10–15 nm.
- Parallel fibers, called intercisternal elements- protein cross-links which hold the cisternae together about 3–6 nm in diameter, existing between the cisternae.
- Several of these cisternae form a functional dictyosome or Golgi field. All dictyosomes in a cell combined to generate the Golgi apparatus.
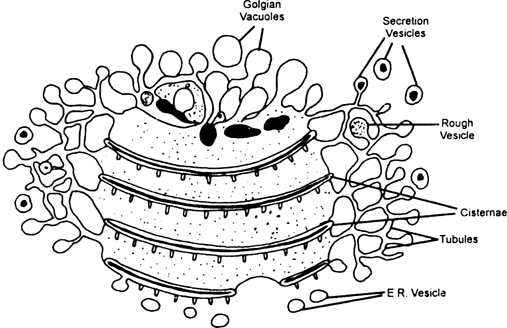
2. Large vacuoles
- These vacuoles are clear and commonly seen at the edge of Complex.
- These structures demonstrate the modified and expanded flattened sacs.
- In the cells of liver and pancreas, these vacuoles of complex include dense masses or granules.
3. Vesicles
Golgi cisternae are always accompanied by Golgi vesicles, which deliver and export material (transport vesicles) and show continuity with cisternae.
- Near the end of distended cisternae are found small spherical vesicles which have probably been pinched off or budding from the distended ends. These are the clusters of dense vesicles that seem to group around the edges of the cisternae.
- Small vesicles 50 nm in diameter remain between the transition elements of the ER and the cis-face of the Golgi stack.
- These structures remain disperse in the surrounding hyaloplasm.
- Investigators suggest that small vesicles associated with the Golgi cisternae fuse with these combined cisternae or fuse with each other to form cisternae of the ER.
Three easily identified vesicle types exist: clathrin-coated vesicles and coat protein complex I and II coated vesicles.
- Clathrin-coated vesicles: They are characterized by a cage-like structure composed of heavy and light clathrin chains that surround the vesicles and primarily bud from the Trans-Golgi Network. They are also engaged in endocytosis from the plasma membrane.
- Coat protein complex I vesicles: They are involved with transport of material from the Golgi to the ER as well as between Golgi stacks.
- Coat protein complex II vesicles: They are associated with the export of secretory material from the ER to the Golgi.
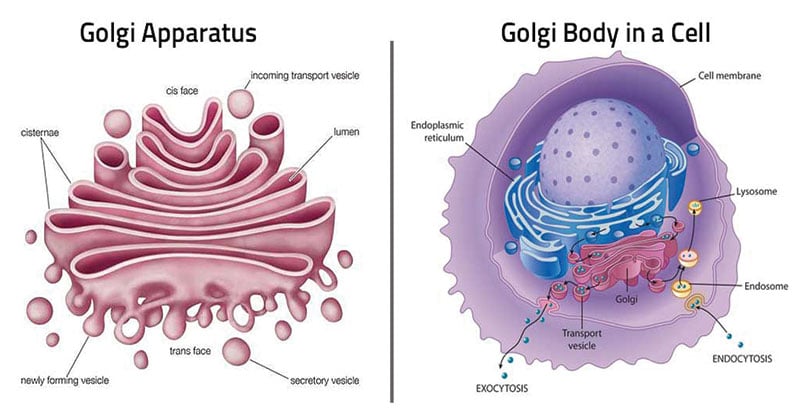
The Golgi apparatus includes three main cisternae as they are defined by their relative position with respect to each other: cis, medial, and trans.
The cis face (convex), is closely associated with the endoplasmic reticulum (ER) or forming face and acts as the receiving center of the Golgi.
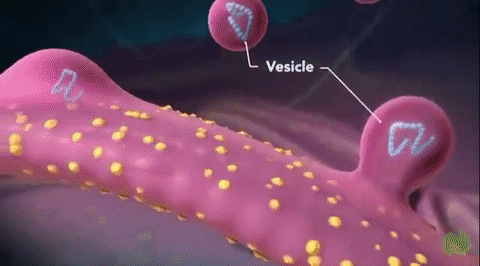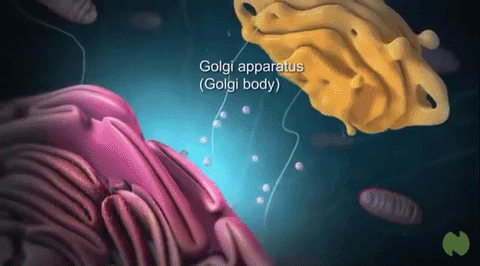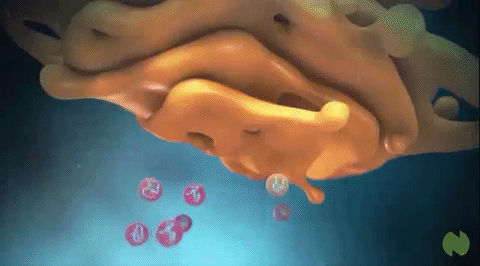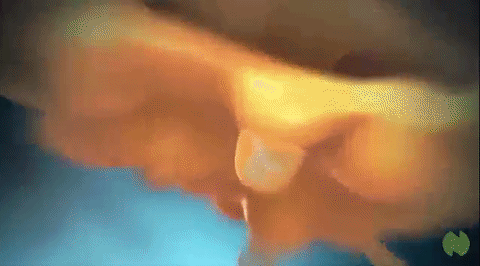
The medial is situated at the middle region of the organelle. At the opposite end of the Golgi stack is a concave trans face or secretory face, which acts as the shipping center from the Golgi apparatus. Trans face is close to the plasma membrane. At the concave side (trans-face) of the cisternae, there are found very small vesicles; among them are coated vesicles, but also somewhat larger vacuoles. They contain variable amounts of secretory products. These two distinct sides also comprise different sets of enzymes.
Functions
There are specific functions that the plant Golgi performs in the cell; it secrets polysaccharides, protein, and lipids, and it serves as a major site for polysaccharide synthesis, protein modification, and glycosylation, or the addition of sugar molecules to proteins and lipids.
- Traffic police of the cell: Dynamics of Golgi trafficking change according to different cellular cues which cause the synthesis and secretion of particular Golgi products. Golgi trafficking changes in relation to the functions of other cellular components, environmental signals, cell stage, and overall needs of the cell. That’s why Golgi vesicles are often, referred to as the “traffic police” of the cell.
-
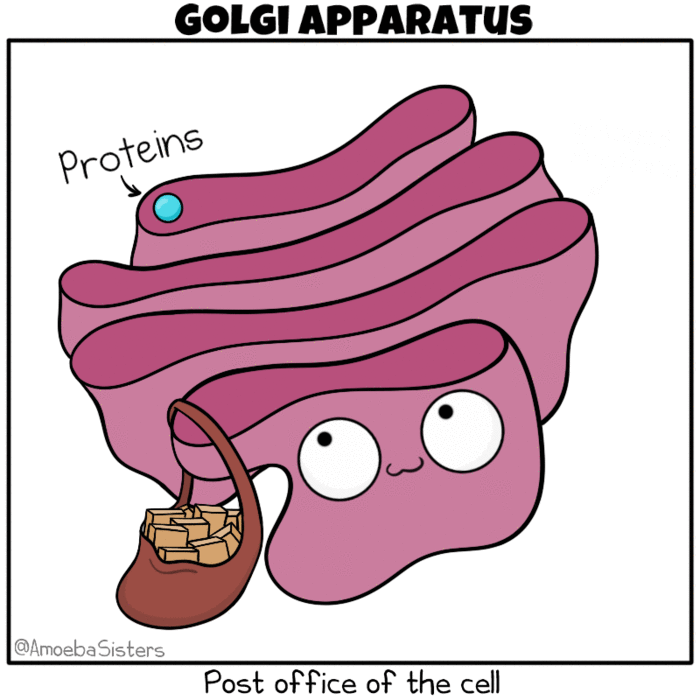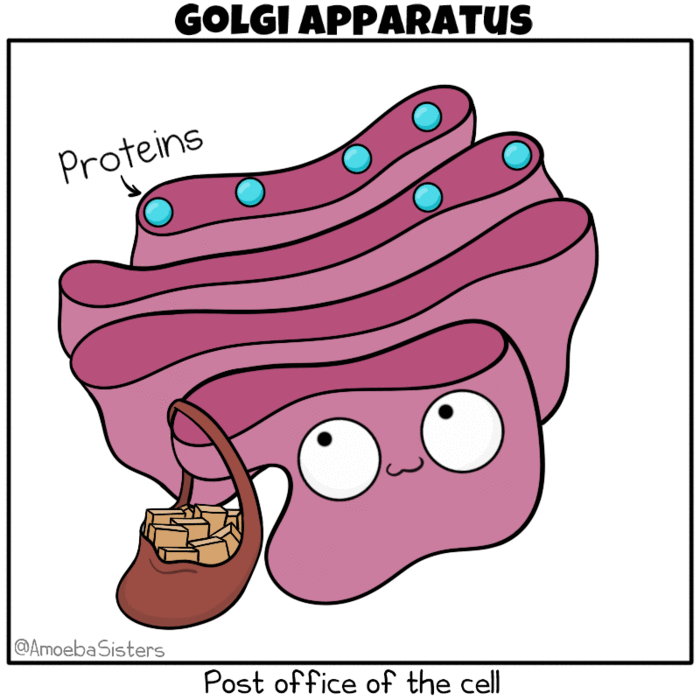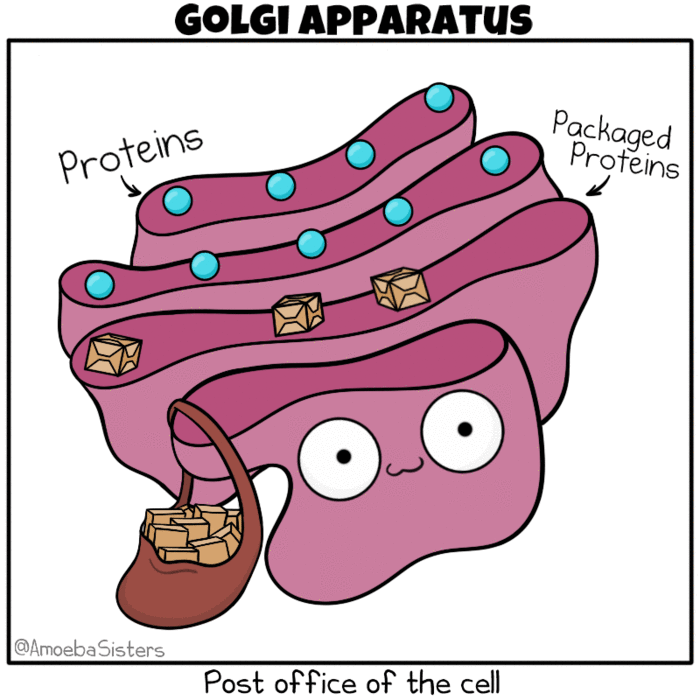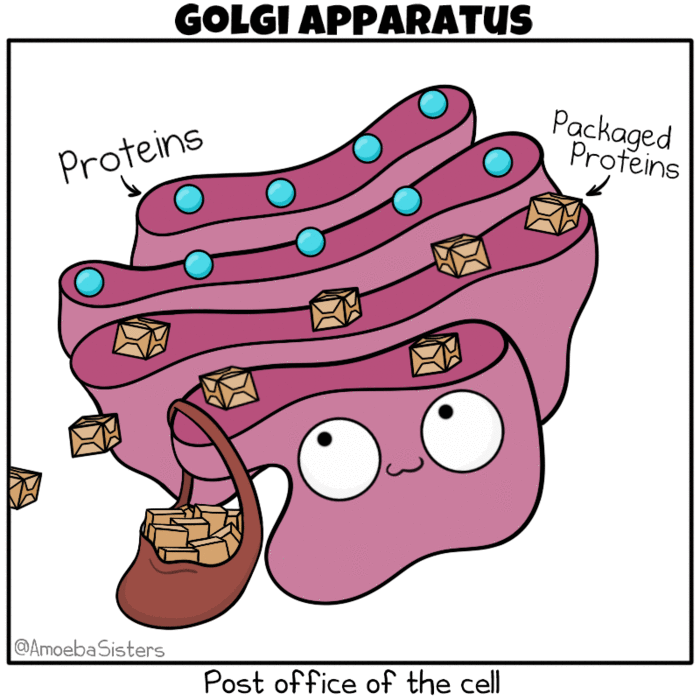
Golgi apparatus as the post office of the cell. Source: The Amoeba Sisters. Modification: Many molecules such as lipids, membrane proteins, and vacuolar enzymes undergo their final modification in the Golgi and then leave it to go to their final destination in the cell. One frequently used analogy describes the ER as a ‘mailbox’ and the Golgi as a ‘post office’, in that it collects and sorts material for shipment from the ER to final destination.
- Formation of secretory vesicles, or cytoplasmic components: The Golgi apparatus is polarized and is asymmetric in both morphologically and functionally. Through transport vesicles that fuse with the Golgi cis face, the complex receives several types of molecules produced in the rough endoplasmic reticulum (RER). After Golgi processing, sequential modification, and packaging, these molecules are released from the Golgi trans face in larger vesicles to constitute secretory vesicles or other cytoplasmic components.
- Storage of Protein: Proteins, which are synthesized in endoplasmic reticulum, migrate to Golgi apparatus, where they are stored and condensed into granular for secretion.
- Formation of Lysosome: Lysosomes may also be produced in the Golgi apparatus.
- Secretion of extracellular materials: In plant and animal cells, the Golgi apparatus is involved in the processing of secretory as well as other glycoproteins. In-plant cells, the Golgi apparatus also participates in the secretion of a variety of extracellular materials, including fucose-rich mucilage by root cap cells, wall-degrading enzymes during abscission, hydrolases that degrade the food reserves during germination, and digestive enzymes in insectivorous plants.
-
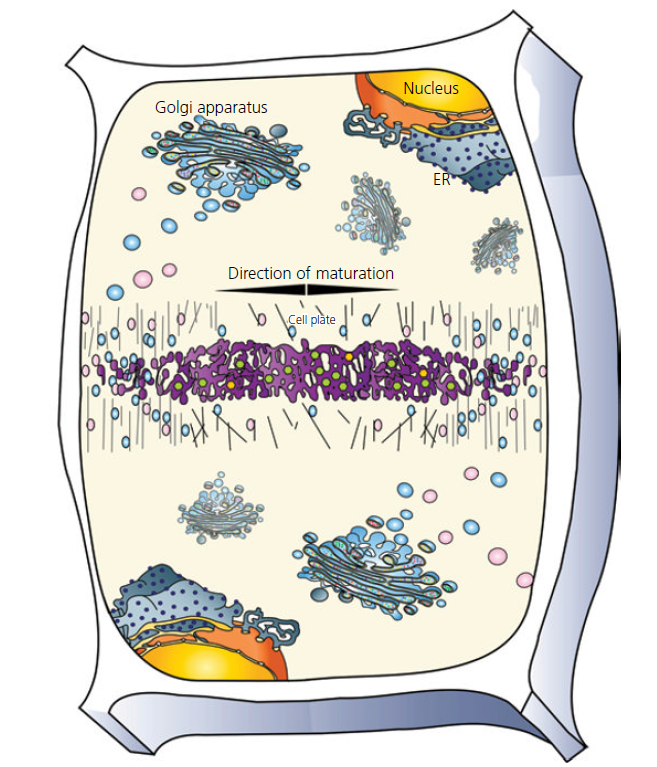
Diagram of a dividing plant cell with multiple intact Golgi actively contributing membranes and polysaccharides to the formation of the cell plate. Source: Plant Cells and Their Organelles by William V. Dashek & Gurbachan S. Miglani. Formation of Cell wall: In plants, the main function of Golgi body is in cell wall formation. Besides modifying newly formed proteins, Golgi bodies also synthesized non-cellulosic cell wall polysaccharides such as hemicellulose and protein. These polysaccharides and glycoproteins are then carried in small vesicles to the plasma- membrane, to the cell plate, where they fuse with the plasmalemma and deposit cell wall materials at the plasmalemma- cell wall interface.
- Synthesis of Carbohydrates: The Golgi apparatus of plant cells secrete complex polysaccharides during cell plate formation and once the cell plate is completed, the Golgi-derived vesicles continue supplying material to the extracellular matrix of the primary- and secondary- cell walls.
- Glycosylations (bound to sugar): Glycosylation, the addition of sugar components, like glycan chains to proteins and lipids, is one of the main functions of the Golgi apparatus. Modification of proteins by adding sugar residues aids in protein stability by preventing degradation as well as affecting protein folding and tertiary structure.
- Transport of Sugars: All sugars must be activated before they are reactive enough to participate in the glycosylation reactions. They become activated after becoming bound to nucleotide diphosphates in the cytosol. Since these are relatively large polar molecules, the membranes of the Golgi apparatus must have the transporters (dolichols or proteins) to transfer the nucleotide-activated sugars into the lumen.
- Processing of Glycoprotein Northcote and Pickett-Heaps (1966) discovered, in radio autographic studies, that the Golgi apparatus is the important site of glucose incorporation in root cells.
- Formation of acrosome: During the maturation of sperm, the Golgi complex plays a role in the formation of acrosome.
- Components of the Golgi apparatus can lessen metal ions.
- The complex activates mitochondria to produce ATP.
Golgi apparatus, being the hub of endomembrane trafficking particularly for the cell wall, vacuole, and other components, has great importance.
Related Articles
References
- Color Atlas of Cytology, Histology and Microscopic Anatomy by Wolfgang Kuehnel.
- Fundamental of Plant Physiology by Dr. V.K. Jain.
- Plant Anatomy by B.P. Pandey.
- Plant Cell Biology from Astronomy to Zoology by Randy Wayne.
- Plant Cells and Their Organelles by William V. Dashek & Gurbachan S. Miglani.
Useful Links


Interesting topic 🧡. Thanks for letting us know 💙🧡
My pleasure
💛