For studying, researching, and observing plants and plant life, we need to start from the beginning of some basic divisions of plants and have the knowledge of some plant categories. Bryophytes are one of the most interesting and incredible divisions of the plant kingdom. Anyone who has the thirst to know about the bryophytes and is curious to know how they reproduce or create generations both in sexual and vegetative ways can have a look at this article.
Let’s get to know what are bryophytes actually? The Bryophytes are like the rest of land plants, Embryophytes (plants that produce an embryo) and have traditionally been viewed as a distinct lineage from other terrestrial plants. They are not considered to have given rise to the vascular plants but they probably were the earliest land plants. Like the rest of the land plants, they evolved from green algal ancestors, closely related to the Charophytes. They are a group of simple land plants, well-adapted to moist habitats.
First of all, let us see how bryophytes reproduce in the sexual (advance) way.
Sexual Reproduction
Sexual reproduction involves the mixing of genes from two different parents to give offspring with a genetic make-up similar to, but different from, each parent.
Best safe and secure cloud storage with password protection
Get Envato Elements, Prime Video, Hotstar and Netflix For Free
Best Money Earning Website 100$ Day
#1 Top ranking article submission website
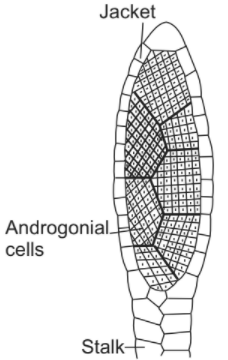
Without exception, the sexual reproduction in all bryophytes is of highly oogamous type, i.e. takes place by motile male gametes (antherozoids) and a large non-motile female gamete (egg). The gametes are produced within the multicellular sex organs, which are provided with an outer sterile layer of jacket. Such an outer layer of the sterile jacket is absent in the sex organs of algae. The male sex organ is called antheridium while the female sex organ is called archegonium.
Antheridium is a multicellular, ellipsoidal, or club-shaped, short-stalked body consisting of a mass of androcytes or antherozoid mother cells surrounded by a single layer of protective sterile jacket cells. Each androcyte produces a single bi-flagellate motile male gamete called antherozoid.
Usually, the antherozoids are spirally coiled bodies, each possessing two whiplash-type flagella. In most bryophytes, the antheridium, on dehiscence, discharges the androcytes in a mass.
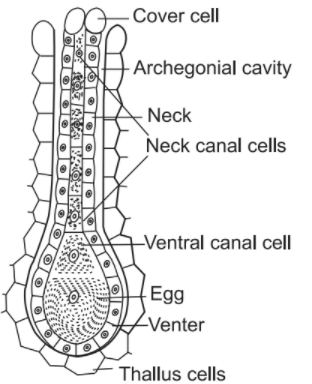
The archegonium is also a multicellular but flask-shaped body with a basal swollen portion called venter and an upper elongated portion called neck. Each archegonium consists of an axial row of cells surrounded by a sterile jacket. The axial row of archegonium contains usually 4–6 or more neck canal cells, a venter canal cell, and a single large basal cell called egg or oosphere. Nourishment and protection to the egg and the developing embryo are provided by the archegonium.
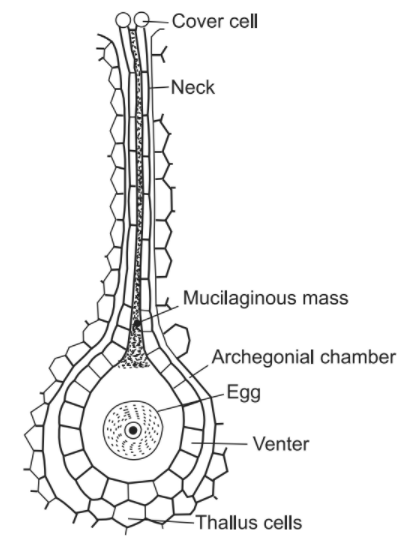
Fertilization takes place only in the presence of water. At the time of fertilization, the axial row of neck canal cells and venter canal cell of the archegonium disintegrate and disorganize forming a mucilaginous liquid. Only the egg remains inside the cavity of the venter. The liberated, flagellated, antherozoids swim in a thin film of water and reach up to the neck of the archegonium. The mouth or the tip of the archegonium also opens at this stage. Several antherozoids pass through the neck canal to the venter, where a single antherozoid fertilizes the egg, and diploid zygote results. Gametes (antherozoids and egg) are the last structures of the gametophytic generation, while the zygote is the first cell of the sporophytic generation.
Vegetative Reproduction
May be defined as a type of asexual reproduction in which a whole new plant is produced from an organ, e.g. a tuber, which is not involved in sexual reproduction. In the absence of mutation, the offspring of the vegetative reproduction will be genetically identical to the parent plant.
Bryophytes reproduce vegetatively by various means. In several of their dioecious species, the plants reproduce mainly by vegetative methods, and some such species have even ceased to reproduce sexually.
1. Death and Decay of Older Parts
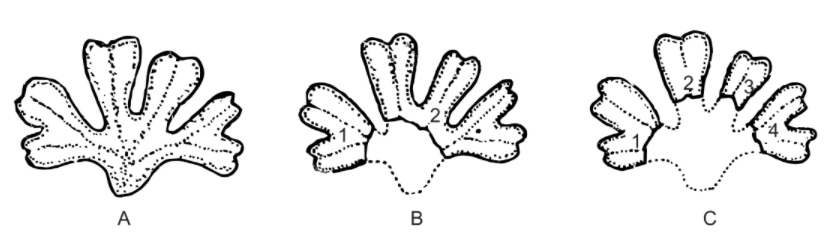
In the majority of Hepaticopsida and Anthocerotopsida, the younger branches of dichotomously dividing thallus get isolated due to progressive death and decay of the older posterior parts. Such isolated parts start functioning as new thalli (Fig. A-C) on return of favourable conditions. This is the most common method of vegetative reproduction in thalloid bryophytes, particularly in liverworts and hornworts. This is also seen in those mosses (Bryopsida) which have prostrate rhizomes bearing erect branches.
2. Adventitious Branches
The term ‘adventitious’ is applied to a plant part developed out of the usual order or in an unusual position. In thalloid bryophytes, adventitious branches usually develop from the underside of the midrib. Upon separation from the parent plant, the adventitious branch develops into a new plant, e.g. Anthoceros laevis, Asterella, Blasia, Corsinia, Dumortiera, Marchantia, Pellia, Reboulia, Riccia fluitans, Sphaerocarpos, Targionia, etc.
3. Innovations
A young offshoot from the stem is known as innovation. On being separated, and falling on the suitable substratum, the innovations develop into new plants, e.g. Sphagnum and many Acrogynous jungermanniales. In Sphagnum, the innovation grows more vigorously than the other branches, continues its upward growth, and takes all the characteristics of the main axis. It is an effective method of multiplication in Sphagnum.
4. Leaf Cladia
A small detachable branch originating from the individual cells of the leaf is known as leaf cladia. On being detached, leaf cladia develop into new plants, e.g. Frullania fragilifolia, Plagiochila, etc.
5. Stem Cladia
It is a small detachable branch which originates from the individual cells of the stem. It occupies almost the same position on the stem as the sex-organ-bearing branch. Stem cladia develops on the stem in several leafy liverworts and mosses, e.g. Bryopteris fruticulosa, Drepanolejeunea, etc.
6. Whole Shoots
All and complete shoots get separated from the parent plant, and on getting suitable water and other requirements develop into new plants, e.g. Pohlia nutans.
7. Shoot Tips
In Campylopus, Polytrichium, and some other mosses, the tips of the shoots get separated and develop into new plants.
8. Modified Branch of Budlike Form
In some species of Bryum and many species of Pohlia and some more mosses, the organ shed is a modified branch of bud-like form. This type of organ is strictly a deciduous branchlet and quite capable of developing into a new plant. In some taxonomic works of bryophytes, such organs have also been termed as bulbil or gemma.
9.Tubers
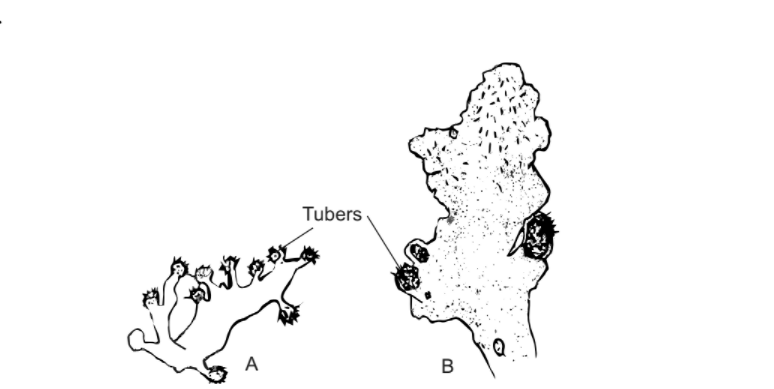
Technically, a tuber is a “thick underground stem in which food is stored”, or, “a swollen underground stem acting as a storage and perennating organ”. Some define tuber as a “swollen part of a stem or root, usually modified for storage”. In bryophytes, tubers develop in several species of liverworts as well as mosses. Some of the common tuber-forming species are Riccia billardieri, R. discolor, Geothallus tuberosus, Asterella angusta, Aitchisoniella himalayensis, Conocephalum conicum, Fossombronia tuberifera, Sewardiella tuberifera, Anthoceros himalayensis and A. laevis
10. Gemmae
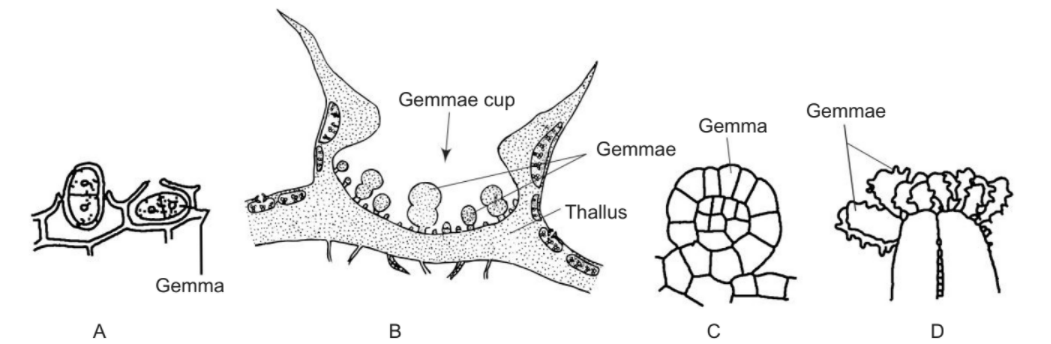
Gemmae (singular: gemma) are “small groups of gr een cells, produced in cup-shaped structures on the surface of thalloid liverworts.” Gemmae are usually dispersed by splashes of rain. Some define gemmae as “a bud that will give rise to a new individual, e.g. the multicellular structure in algae, pteridophytes and specially bryophytes”. These are the means of vegetative propagation and form abundantly in liverworts (Hepaticopsida), to some extent in hornworts (Anthocerotopsida) and to a lesser extent in mosses (Bryopsida). Gemmae are not formed in Sphagnales. Gemmae reported in different groups of bryophytes are listed below.
Gemmae of Liverworts (Hepaticopsida): Many types of gemmae are produced in Hepaticopsida. Of these, some are listed below:
- One to three-celled gemmae developing on the stem apex, e.g. Cephalozia bicuspidata, Lophozia heterocolpa.
- One- to three-celled gemmae developing on the leaves, e.g. Cephalozia francisci, Lophozia barbata.
- Two-celled endogenous gemmae developing within any external cell of the thallus, e.g.
Riccardia multifida , Haplozia caespiticia. - Three- to four-celled gemmae developing in the axils of the leaves, e.g. Treubia.
- Stalked, multicellular, discoid gemmae formed on the dorsal surface of the thallus inside gemma cups, e.g. Marchantia (Fig. B), Lunularia. (vi) Subspherical gemmae produced in large number in flask-shaped gemma receptacles e.g. Blasia.
- Star-shaped gemmae developing on the dorsal surface of the thallus, e.g. some species of
Blasia. - Discoid, multicellular gemmae developing on leaves, e.g. Rudula complanata (Fig. C),
Leptocolea. - Discoid multicellular gemmae developing on erect gemmiferous branches, e.g. Metzgeria uncigera (Fig. D).
Gemmae of Hornworts (Anthocerotopsida): Some Anthoceros species bear gemmae. In A. glandulosus, several gemmae develop along the margin of the thallus, and a few also develop on the dorsal surface. Gemmae have also been reported in some more species, such as A. formosae and A. propaguliferus.
Gemmae of Mosses (Bryopsida): Six types of gemmae developing on different plant parts of mosses are listed as under:
- On Rhizoids of Leafy Shoots These are the multicellular gemmae developing on the rhizoids of leafy shoots, e.g. Barbula convoluta, Bryum erythrocarpum, etc.
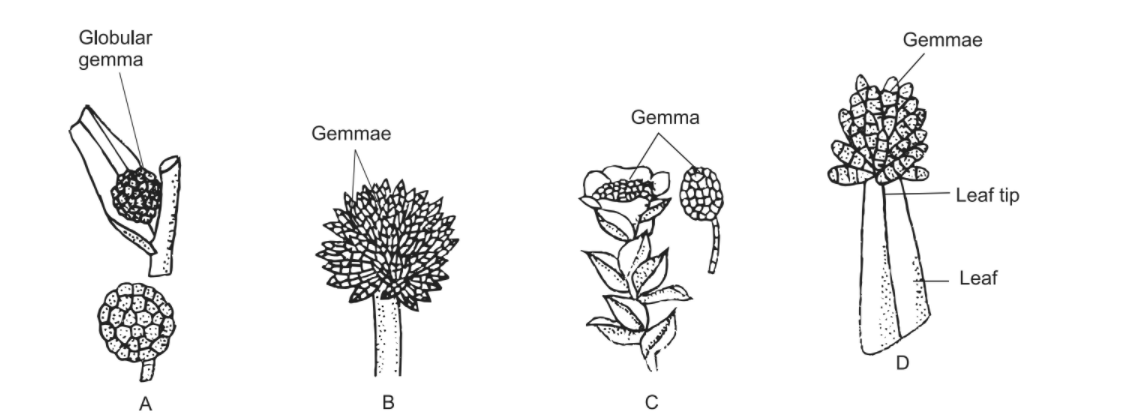
- At the Base of Stem In Bryum erythrocarpum (Fig.A) and B. rubens, multicellular and globular gemmae develop at the base of the stem.
- At the End of Leafless Stalks In Aulacomnium androgynum (Fig.B), stalked fusiform gemmae develop at the end of leafless stalks.
- At the Tip of the Shoot In Tetraphis pellucida (Fig.C), green stalked, multicellular and lenticular gemmae develop at the tip of shoot. Widened leaves form a cup-like structure around such gemmae.
- On the Stem and Branches In Pterygynandrum filiforme, , smooth, golden-brown gemmae develop on the stems and branches. Such gemmae are ovoid and stalked bodies. These are bi-celled structures and develop on colorless stalks made of three or more cells.
- On the Leaves In Torula papillosa, Ulota phyllantha (Fig.D) and some more bryophytes, multicellular, articulate gemmae develop on the leaves.
11. Primary Protonema
In almost all mosses, the spore germinates into a young filamentous and branched gametophyte, known as primary protonema. It usually breaks into smaller parts or fragments, and each such fragment is capable of developing into a new protonema and forms gametophyte initials.
12. Secondary Protonema
A protonema-like structure, developing by other methods than from the germination of spores is known as secondary protonema. It is also a means of vegetative reproduction in Bryopsida (e.g. Funaria hygrometrica). Secondary protonema develops (i) either from the rhizoids of a leafy gametophore when exposed to light, or (ii) from almost any separated living part of the moss plant such as stem, leaves, sex organs, or even paraphysis and sterile cells of seta and capsule.
References
- Bryophyta by O. P. Sharma
Revised by
- Khaleda Akter Shompa on 6 September, 2021